Несколько месяцев назад был опубликован цикл статей под общим названием «Логика мышления». Оптимистично планировалось продолжить его через месяц-другой. Но жизнь внесла свои коррективы. Моделирование паттерно-волновой модели коры дало настолько интересные результаты, что пришлось на время отложить все остальное, в том числе и написание продолжения цикла для хабра.
Однако, не так давно я написал и выложил на препринт статью (http://arxiv.org/abs/1406.6901). В чем-то она может быть интересна тем, кто ранее заинтересовался волновой моделью. Напомню, что ключевой момент модели – это утверждение, что нейроны способны запоминать и узнавать не один единственный образ, описываемый весами его синапсов, а еще и огромное количество других отличных от этого образа сигналов. Конечно, такое усложнение нейрона идет в разрез со многими существующими теориями и требует более, чем серьезного обоснования. Ниже я, как раз, и попробую описать один из приведенных в статье аргументов в пользу моей модели.
Эту статью не стоит воспринимать как продолжение цикла, она скорее приквел к нему. Для себя я назвал это рассуждениями о ключевой роли дендритных шипиков.
Начнем с повторения хорошо известного.
В состоянии покоя между внутренней и внешней средой нейрона существует разность потенциалов – мембранный потенциал, составляющий около 70 милливольт. Он образуется за счет белковых молекул, работающих как ионные насосы. В результате мембрана приобретает поляризацию, при которой отрицательный заряд накапливается внутри клетки, а положительный снаружи.
Поверхность нейрона покрыта ветвящимися отростками – дендритами. К телу нейрона и к его дендритам примыкают аксонные окончания других нейронов. Места их соединений называются синапсами. Посредством синаптического взаимодействия нейрон способен реагировать на поступающие сигналы и при определенных обстоятельствах генерировать собственный импульс, называемый спайком.
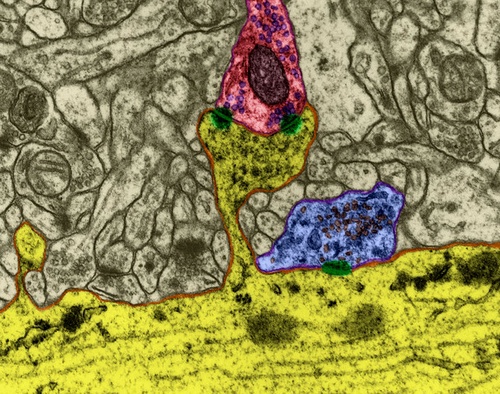
Передача сигнала в синапсах происходит за счет выделения нейромедиаторов. Когда нервный импульс по аксону поступает в пресинаптическую терминаль, он высвобождает из синаптических пузырьков молекулы нейромедиаторов, характерные для этого синапса. На мембране нейрона, получающего сигнал, располагаются рецепторы, которые взаимодействуют с нейромедиаторами.
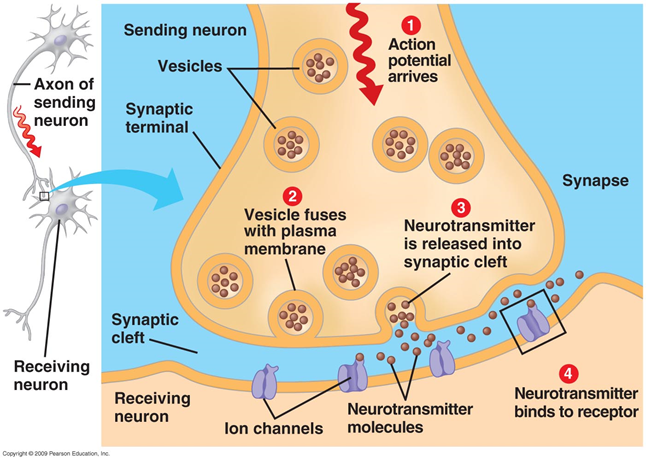
Рисунок 1. Химический синапс
Рецепторы, расположенные в синаптической щели, по большей части являются ионотропными. То есть они же являются ионными каналами, способными перемещать ионы через мембрану нейрона. Нейромедиаторы так воздействуют на рецепторы, что их ионные каналы открываются. В результате мембрана либо деполяризуется, либо гиперполяризуется – в зависимости от того, какие каналы затронуты и, соответственно, какого типа этот синапс. В возбуждающих синапсах открываются каналы, преимущественно пропускающие катионы внутрь клетки, – мембрана деполяризуется. В тормозных синапсах открываются каналы, выводящие катионы из клетки, что приводит к гиперполяризации мембраны.
В определенных обстоятельствах синапсы могут менять свою чувствительность, что называется синаптической пластичностью. Это приводит к тому, что одни синапсы становятся более, а другие менее восприимчивы к внешним сигналам.
Одновременно на синапсы нейрона поступает множество сигналов. Тормозящие синапсы смещают потенциал мембраны в сторону накопления заряда внутри клети. Активирующие синапсы, наоборот, стараются разрядить нейрон. Когда суммарная деполяризация превышает порог инициации, возникает разряд, называемый потенциалом действия или спайком.
После выброса нейромедиаторов специальные механизмы обеспечивают их утилизацию и обратный захват, что приводит к расчистке синаптической щели и окружающего синапс пространства. В течение рефрактерного периода, наступающего после спайка, нейрон не способен порождать новые импульсы. Продолжительность этого периода определяет максимальную частоту генерации, на которую способен нейрон.
Теперь опишем факты менее известные.
Когда потенциал действия, распространяясь по аксону, доходит до нейрона получателя, он вызывают выброс нейромедиаторов в синаптическую щель. Эти медиаторы определяют вклад синапса в общее изменение мембранного потенциала нейрона, получающего сигнал. Но часть медиаторов попадает за пределы синаптической щели и растекается по пространству, образованному нейронами и окружающими их глиальными клетками. Это явление называется спиловером (spillover (анг.) – переполнение, перелив) (Kullmann, 2000). Кроме того, медиаторы испускаются несинаптическими терминалями аксонов и глиальными клетками (Рисунок 2). Концентрация нейромедиаторов вне синапсов оказывается много меньше, чем в синаптических щелях. Однако именно в этих «растекшихся» нейромедиаторах таится масса интересного.
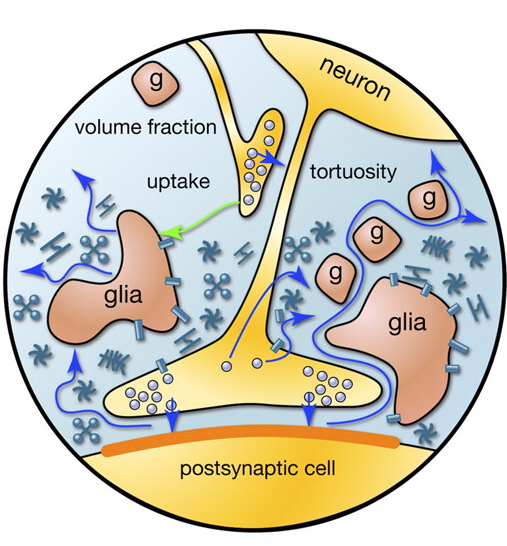
Рисунок 2. Источники медиаторов за пределами синаптической щели (Sykova E., Mazel T., Vagrova L., Vorisek I., Prokopova-Kubinova S., 2000)
Попробуем оценить количество и структуру источников, выбрасывающих нейромедиаторы за пределы синапсов. Для этого воспользуемся количественными оценками параметров коры, приведенными в таблице ниже (Таблица 1) (Braitenberg V., Schuz A., 1998).
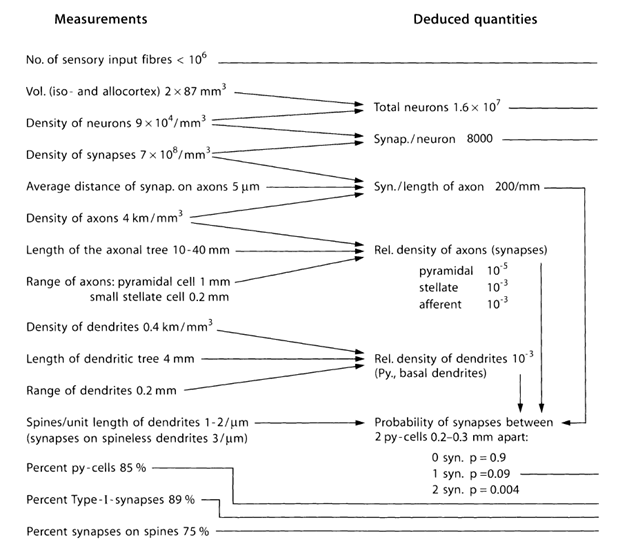
Таблица 1. Сводная таблица параметров, полученная при изучении мозга мыши (py-cell – пирамидальная клетка, Type-I – синапсы между двумя пирамидальными клетками) (Braitenberg V., Schuz A., 1998)
Напомню, что большая часть синапсов (90-95%) приходится не на тело нейрона, а на его дендриты. Дендриты – это тонкие ветвящиеся отростки, образующие так называемое дендритное дерево нейрона. На рисунках ниже дендритные деревья выделены черным, а аксоны серым. Для нейронов разных типов формы дендритных деревьев различны, но сохраняется общий принцип: дендритное дерево состоит из множества ветвящихся отростков, при этом наибольшая плотность синаптических соединений нейрона приходится на небольшую пространственную область. Для основных типов нейронов она составляет порядка 200 мкм (Рисунок 3, Рисунок 4).

Рисунок 3. Структура звездчатого нейрона, линейка – 0.1 мм (Braitenberg, 1978)
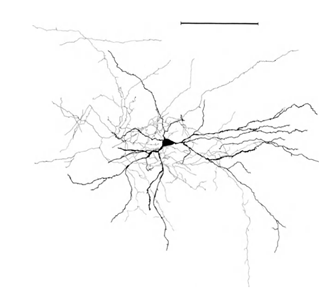
Рисунок 4. Структура пирамидального нейрона, линейка – 0.1 мм (Braitenberg, 1978)
Ветвящиеся аксоны нейронов образуют контакты (синапсы) с дендритами других нейронов. Среднее расстояние между синапсами на дендритах – 0.5 микрометра. Среднее расстояние между синапсами на аксонах – 5 микрометров, то есть в 10 раз больше. Не удивительно, что аксоны получаются приблизительно в 10 раз длиннее чем дендриты.
Большая часть (75 %) синапсов располагается на дендритных шипиках, что наиболее характерно для пирамидальных клеток (Рисунок 5).
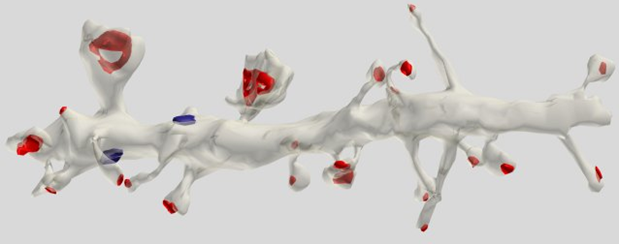
Рисунок 5. Сегмент дендрита пирамидальной клетки. Красным отмечены синапсы на шипиках, синим – на дендритном стволе (Dr. Kristen M. Harris)
Компьютерное моделирование, выполненное на основе реальных анатомических и физиологических данных, показало, что, например, глутамат может распространяться за пределы синаптической щели в количествах, достаточных для активации NMDA рецепторов в радиусе, сопоставимом с расстоянием между соседними синапсами (0.5 мкм) (Rusakov D. A., Kullmann D. M., 1998). Можно предположить, что существенная концентрация нейромедиаторов после спиловера наблюдается на участке дендрита длиной порядка 1-2 мкм. На таком участке может располагаться порядка двух-четырех синапсов, принадлежащих этому дендриту.
Если взять участок дендрита длиною 5 мкм (Рисунок 6), то ожидаемое число синапсов на нем составит около 10.
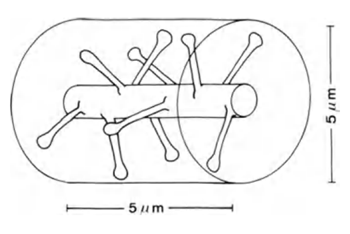
Рисунок 6. Участок дендрита (Braitenberg V., Schuz A., 1998)
Но дендритные ветки одних нейронов тесно переплетаются с ветками других нейронов. Они проходят друг от друга в самой непосредственной близости. За счет высоты дендритных шипиков синапсы, принадлежащие одному дендриту, могут оказаться ближе к поверхности другого дендрита, чем его собственные синапсы.
Если бы синапсы были равномерно распределены в пространстве коры, то в цилиндрический объем высотою 5 мкм и диаметром тоже 5 мкм (рисунок выше) при плотности распределения синапсов 7x108/мм3 попало бы около 100 синапсов. То есть в 10 раз больше того, что непосредственно располагается на самой ветке. В действительности существенную часть объема мозга занимают глиальные клетки и тела нейронов, что еще больше повышает плотность упаковки синапсов. Однако, нейроны работают с разными нейромедиаторами, что тоже надо учитывать.
Теперь попробуем понять смысл такой структуры связей с точки зрения распределения плотности внесинаптического медиатора. Для этого воспользуемся упрощенной моделью. Возьмем условный окружающий нейрон объем и занумеруем входящие в него нейроны. Каждый из этих нейронов будет иметь:
• несколько синаптических контактов с дендритом выбранного нейрона;
• несколько «прилеганий», то есть мест, где его синапсы с другими нейронами будут располагаться в непосредственной близости от дендрита выбранного нейрона.
Представим дендритное дерево как одну длинную ветку с равномерно распределенными условными источниками (Рисунок 7). Для каждого источника на этой ветке можно указать номер нейрона из окружающего пространства, отвечающего за него. Каждый из нейронов окружения будет иметь сразу несколько источниковых контактов, случайным образом распределенных по дендриту. Обозначим это соотнесение вектором D с элементами di.
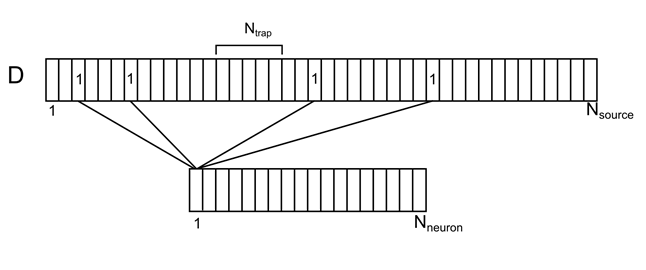
Рисунок 7. Соотнесение окружающих нейронов и их контактов на дендрите
Введем обозначения:
N neuron – количество нейронов окружения
N source – количество источников для одного нейрона
N trap – количество источников, создающих уровень плотности нейромедиаторов (синаптическую ловушку)
Теперь предположим, что несколько нейронов из окружения дали спайк. Это можно воспринимать как сигнал, доступный для наблюдения нашему нейрону. Обозначим N sig– количество активных нейронов, создающих информационный сигнал. Запишем этот сигнал бинарным вектором S.
Для всех позиций на дендрите кроме самых крайних будем считать плотность медиатора по формуле
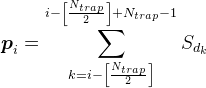
Например, для сигнала, показанного на рисунке ниже, плотность в отмеченной синаптической ловушке составит 2 (сумма сигналов от 1-го и 4-го нейронов).
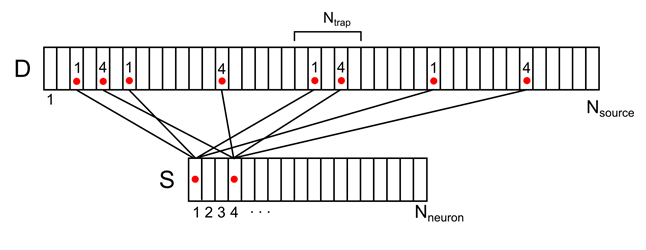
Рисунок 8. Отображение активности двух нейронов окружения на дендритное дерево (показана только часть связей и нумерации)
Для любого произвольного сигнала можно рассчитать картину плотности медиаторов на дендрите. Эта плотность будет колебаться в диапазоне от 0 до N trap. Максимальное значение будет достигаться, если оказались активны все источники, образующие синаптическую ловушку.
Воспользуемся усредненными значениями параметров, характерных для реальной коры крысы (Braitenberg V., Schuz A., 1998). Исходя из них мы получим следующие параметры модели:
N neuron = 650
N source = 25000
N trap = 15
Будем полагать, что сигнал кодируется активностью, например, 1.5% нейронов коры, тогда
N sig = 10
Несложно рассчитать вероятность того, что для произвольного сигнала, состоящего из N sig единиц, найдется хотя бы одно место на дендрите, где плотность медиатора составит ровно K. Для приведенных параметров вероятность принимает следующие значения (Таблица 2):
| K | P |
| 0 | 0,984 |
| 1 | 1 |
| 2 | 1 |
| 3 | 0,996 |
| 4 | 0,287 |
| 5 | 0,016 |
| 6 | 0,001 |
| 7 | 0 |
| 8 | 0 |
| 9 | 0 |
| 10 | 0 |
| ... |
Таблица 2. Таблица вероятности нахождения хотя бы одной ловушки с заданной плотностью. Первый столбец – требуемое количество активных источников в ловушке. Второй – вероятность нахождения на дендрите хотя бы одного места, где будет именно такое количество активных источников
То есть при тех значениях параметров, что близки к конфигурации реальной коры, для любого объемного сигнала, затрагивающего около 1.5% нейронов, справедливо следующее:
• Найдется около 1.6% нейронов, у которых на дендритах существует ловушка, где пересекается 50% сигнальных аксонов;
• Практически у каждого нейрона найдутся ловушки, в которых пересекается хотя бы 30% сигнальных аксонов.
Смысл такого результата очень интересен. Предположим, что информация в коре каким-то образом кодируется синхронной активностью относительно небольшого количества (N sig) компактно расположенных нейронов. Речь идет не обо всей деятельности мозга, а об информационных процессах в малом объеме, где нейроны занумерованы нами от 1 до N neuron. Предположим, что количество кодовых комбинаций S ограничено и образует определенный словарь T емкостью N dict. Можно рассчитать вероятность того, что одно и тоже место «откликнется» сразу на два сигнала. Результаты такого расчета для словаря из 10000 сигналов в таблице ниже.
| K | P error |
| 3 | 0,00399 |
| 4 | 1,05E-05 |
| 5 | 1,89E-08 |
| 6 | 2,33E-11 |
| 7 | 0 |
| ... |
Таблица 3. Таблица вероятности нарушения уникальности синаптической ловушки при различных уровнях плотности медиатора
Получается, что при K=3 ловушки обладают определенной селективностью, хотя и не гарантируют от ошибки, но уже при K=5 они начинают достаточно однозначно соответствовать определенному пространственному паттерну активности. Напомню, что это справедливо не для произвольных сигналов из бесконечного набора, а когда мы имеем набор, пусть и достаточно большой, дискретных разрешенных состояний активности.
То есть получается, что смысл свойственной мозгу структуры аксонных и дендритных деревьев заключается в создании на дендритной поверхности каждого нейрона богатого пространства ловушек, то есть мест, соответствующих всевозможным комбинациям источников нейромедиаторов. По плотности нейромедиатора в ловушках можно судить о пространственно распределенном сигнале, складывающемся из синхронной активности некоторого числа близлежащих нейронов.
А так как на поверхности нейрона располагаются так называемые метаботропные рецепторы, которые могут вызывать одиночные спайки нейрона даже при незначительной концентрации нейромедиаторов, то выходит, что этими спайками, которые принято называть спонтанными, нейрон может реагировать на огромное количество разных сигналов, отличных от тех на которые могут быть настроены синапсы нейрона.
Собственно, теперь можно объяснить назначение тех самых дендритных шипиков с которых начиналась статья. По идее, все синапсы могли бы располагаться непосредственно на дендрите или теле нейрона. Это никак не повлияло бы на прямую работу синапсов и их способность давать вклад в появление вызванной активности. Но теперь можно предположить, что назначение дендритных шипиков – это создание пространственной структуры, в которой синапсы разных нейронов оказываются «перемешаны» так, что приобретают способность своим спиловером влиять не только на поверхность своего, но и окружающих соседних дендритов. Такая вот получается «принципиальная мелочь».
В самой исходной статье показывается как из этого всего рождается собственно «волновая история», но это уже отдельный разговор.
И в завершении о небольшой корысти. Исходная статья в оригинале на русском. Есть перевод на английский язык, сделанный Дмитрием Шабановым (за что ему огромный респект), но он (перевод) далек от совершенства. Если у кого есть возможность посмотреть и указать на ляпы буду крайне признателен. Документ по ссылке открыт всем для комментирования.
Список литературы к исходной статье
1. (2014). Получено из Human Connectome Project: www.humanconnectomeproject.org
2. (2014). Получено из ALLEN Mouse Brain Connectivity Atlas: connectivity.brain-map.org
3. Bloom, B. H. (1970). Space/time trade-offs in hash coding with allowable errors. Communications of the ACM Т. 13 (7), 422–426.
4. Braitenberg V., Schuz A. (1998). Cortex: statistics and geometry of neuronal connectivity, 2nd ed.
5. Braitenberg, V. (1978). Cortical architectonics: general and areal. В M.A.B. Brazier and H. Petsche (eds), Architectonics of the Cerebral Cortex (стр. 443–465). New York: Raven Press.
6. Coster, H. (1975). Electromechanical stresses and the effect of pH on membrane structure. Biochim Biophys Acta 13;382(2), 142-146.
7. Dr. Kristen M. Harris. (б.д.). Synapse Web. Получено из synapses.clm.utexas.edu.
8. Fukushima, K. (1980). Neocognitron A self-organizing neural network model for a mechanism of pattern recognition unaffected by shift in position. Biological Cybernetics, 36(4), 193-202.
9. Grossberg, S. (1987). Competitive learning: From interactive activation to adaptive resonance. Cognitive Science N11, 23-63.
10. Hebb, D. (1949). The Organization of Behavior. New York: John Wiley & Sons.
11. Hodgkin, A. a. (1952). A quantitative description of membrane current and its application to conduction and excitation in nerve… J. Physio l. 117, 500—544.
12. Izhikevich, E. M. (2007). Dynamical Systems in Neuroscience: The Geometry of Excitability and Bursting. London: The MIT Press.
13. Kullmann, D. M. (2000). Spillover and synaptic cross talk mediated by glutamate and GABA in the mammalian brain. Prog Brain Res, 125, 339-351.
14. Kuramoto, Y. (1984). Chemical Oscillations, Waves, and Turbulence. Dover Publications.
15. Liibke J., Markram H., Frotscher M., Sakmann B. (1996). Frequency and dendritic distribution bution of autapses established by layer 5 pyramidal neurons in the developing rat neocortex: comparisons with synaptic innervation of adjacent neurons of the same class. Neurosci 16 (10), 3209-3218.
16. Malenka R.C., Nicoll R.A. (1999). Long-term potentiation--a decade of progress? Science 285(5435), 1870-1874.
17. Michael T. Lippert, Kentaroh Takagaki, Weifeng Xu, Xiaoying Huang, Jian-Young Wu. (2007). Methods for Voltage-Sensitive Dye Imaging of Rat Cortical Activity With. J Neurophysiol 98, 502-512.
18. Pitts W., McCulloch W.S. (1947). How we know universals: the perception of auditory and visual forms. Bull. Math. Biophys V.9, 127—147.
19. Rosenblatt, F. (1962). Principles of Neurodynamic: Perceptrons and the Theory of Brain Mechanisms.
20. Rusakov D. A., Kullmann D. M. (1998). Extrasynaptic glutamate diffusion in the hippocampus: ultrastructural constraints, uptake, and receptor activation. Neurosci 18(9), 3158-3170.
21. Sheng, M., Nakagawa, T. (2002). Neurobiology: glutamate receptors on the move. Nature, 417(6889), 601-602.
22. Sheng, M., Sala C. (2001). PDZ domains and the organization of supramolecular complexes. Annu Rev Neurosci, 24, 1-29.
23. Sykova E., Mazel T., Vagrova L., Vorisek I., Prokopova-Kubinova S. (2000). Extracellular space diffusion and pathological states. Progress in Brain Research, 155-178.
24. Tovar K. R.,Westbrook G. L. (2002). Mobile NMDA receptors at hippocampal synapses. Neuron, 34(2), 255-264.
25. W.-F. Xu, X.-Y. Huang, K. Takagaki, and J.-Y. Wu. (2007). Compression and reflection of visually evoked cortical waves. Neuron, 55, 119-129.
26. Y. LeCun, Y. Bengio. (1995). Convolutional Networks for Images, Speech, and Time-Series, in Arbib, M. A., editor, The Handbook of Brain Theory and Neural Networks. MIT Press.
27. Ж.П. Шнурова, З.М. Гвоздикова. (1971). Реакция нейронов сенсомоторной области коры на ее прямое электрическое раздражение. В Сборник «Исследование организации нейронной деятельности коры больших полушарий головного мозга» (стр. 158-180). Москва: Наука.
28. Николлс Дж., Мартин Р., Валлас Б., Фукс П. (2003). От нерона к мозгу (четвертое издание).
29. Прибрам, К. (1971). Языки мозга.
30. Радченко, А. Н. (2007). Информационные механизмы мозга. Спб.
31. Редозубов, А. (2014). Программа для расчета распределения синаптических ловушек. Получено из aboutbrain.ru: www.aboutbrain.ru/programs
32. Редозубов, А. (2014). Программа моделирования волновой коры. Получено из aboutbrain.ru: www.aboutbrain.ru/programs








